Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2009/2013
[A propos de l'auteur]
[Droits de copie]
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2009/2013
[A propos de l'auteur]
[Droits de copie]
On suppose un bioréacteur alimenté par un apport de bulles d'un gaz contenant du dioxygène. On suppose que les 2 phases "bulles de gaz" et milieu liquide de culture sont parfaitement homogènes à l'intérieur du bioréacteur. On suppose que le réacteur est suffisament peu profond pour négliger les différences de pression entre le fond et la surface !
Dans le bioréacteur, du dioxygène est transféré grâce au système d'aération (bullage) et du dioxygène est consommé par la biomasse. On peut alors écrire les relations fondamentales :
Transfert de dioxygène | Consommation de dioxygène |
Soit OTR la vitesse de transfert de l'O2 dans le réacteur (Oxygen Transfer Rate) |
Soit OUR la vitesse de consommation de O2 par la biomasse (Oxygen uptake Rate) |
OTR = KLa(C*-C) (1)
• OTR est exprimé en quantité d'O2
par unité de volume et par unité de temps (mol.m-3s-1)
• C : concentration en O2 dissous dans le milieu liquide de culture supposé parfaitement homogénéisé (mol/L) (1) signifie très simplement que le transfert de dioxygène est proportionnel à un coefficient égal au produit du coefficient KL et du coefficient "a" (et on va pouvoir contrôler la facteur "a", la surface d'échange air/milieu) et proportionnel au gradient de dioxygène C*-C (et on pourra avoir intérêt à améliorer ce facteur par un C* élevé en jouant éventuellement sur la pression et la richesse en dioxygène du gaz d'aération). |
OUR = QO2 X (2)
• OUR est exprimé en quantité d'O2
par unité de volume et par unité de temps (mol.m-3s-1)
• QO2 : vitesse spécifique de consommation du dioxygène par la biomasse (masse de O2 par unité de masse de biomasse et par unité de temps,
g O2 . g-1 X . h -1) (2) montre que la consommmation est proportionellement plus élevée si la biomasse est plus élevée ... assez trivial. |
|
(1) et (2) conduisent alors très simplement à la relation bilan dynamique fondamentale : dC/dt = OTR - OUR
= KLa(C*-C)
- QO2 X
|
|
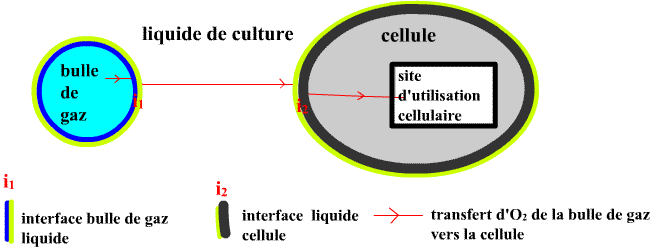
Le transfert d'O2 d'une bulle gazeuse vers un site d'utilisation dans un microorganisme comporte plusieurs
étapes :
• Passage de la bulle de gaz vers le milieu liquide supposé parfaitement homogénéisé : 2 étapes : passage à travers le film gazeux et le film liquide constituant l'interface
gaz-liquide.
• Diffusion/convection dans le milieu de culture liquide supposé parfaitement homogénéisé.
• Passage à travers l'interface milieu de culture-cellule.
• Diffusion vers le site d'utilisation. Pas simple chez les eucayotes par exemple où l'intervention finale de
O2 est mitochondriale ...
Evidemment tout ça se théorise avec de belles équations ! (on peut même y introduire des contacts plus ou moins directs bulles de gaz/cellules aux fortes concentrations en microorganismes ... etc ... etc .... ). Mais ceci ne sera pas le prospos de ces pages. Néanmoins, vous pouvez ouvrir une nouvelle fenêtre qui vous en dira un peu plus en cliquant sur ce lien.
Il faut retenir que c'est l'étape de passage du dioxygène à travers le film liquide de l'interface bulle de gaz/liquide qui est limitante des capacités de transfert du dioxygène (la première étape i1 de l'ensemble des étapes proposées ci-dessus).
Quoiqu'il en soit, les deux petits tableaux ci-dessous mériteront quelques minutes d'attention.
Commentaire |
Grandeur, unité |
|
KL |
KL a la dimension d'une vitesse de parcours et se rapporte à l'unité de surface de l'interface gaz-liquide. |
D'où sa grandeur en longueur.temps-1 |
Coef. "a" |
Le coefficient "a" représente la surface d'interface gaz-liquide par unité de volume de bioréaction. |
D'où sa grandeur en surface.volume-1 |
KLa |
Le coefficient "a" est généralement inconnu, et seul le produit KLa est accessible. On appelle coefficient volumétrique de transfert le produit KLa. |
En faisant le produit des dimensions de KL et de "a", on trouvera la dimension de KLa, un temps-1. |
KLa(C*-C) |
Et il n'est pas inutile de faire observer au lecteur que KLa(C*-C) a bien pour dimension un temps-1 multiplié par une concentration, c'est à dire un débit de concentration (un flux volumétrique de dioxygène). Ouf, tout va bien ! Et la relation OTR = KLa(C*-C) traduit tout simplement le fait que la vitesse de transfert du dioxygène est limitée
par le passage bulle de gaz/milieu liquide et dépend ainsi de façon linéaire de
3 facteurs : Et un peu de bon sens. Le coef. "a" (surface d'interface gaz-liquide par unité de volume) conduit à l'évidence suivante : plus les bulles seront petites et nombreuses, plus "a" sera grand et meilleur sera le transfert de dioxygène ! On s'en doutait un peu ... |
Quantité par unité de temps et par unité de volume, quantité.volume-1temps-1.
|
Conditions | Commentaire |
|
Mesures de KLa en absence d'activité respiratoire (OUR = 0) |
On peut mesurer KLa en absence de microorganismes dans le bioréacteur, en travaillant avec le seul milieu de culture dans le fermenteur. On peut aussi, dans les mêmes conditions de milieu, travailler en présence de microorganismes mais avec un artifice bloquant la respiration. On a alors OUR = 0. |
|
Mesures de KLa en présence de microorganismes en respiration |
Aux concentrations pas trop élevées en microorganismes les KLa mesurés en absence d'activité respiratoire ou en présence de microorganismes en respiration sont identiques (toutes choses égales par ailleurs) : c'est bien l'étape de passage du dioxygène à travers le film liquide de l'interface bulle de gaz/liquide qui est limitante des capacités de transfert du dioxygène. Et pas la suite du transfert : le passage à travers le film de contact milieu/cellules puis le passage au site de consommation. En revanche, les fortes activités respiratoires vont entraîner un facteur d'amplification de KLa. Ce facteur est généralement appelé facteur d'amplification biologique E (biological enhancement factor). |
|